23 March 2022: Laboratory Research
Production of Inflammatory Mediators in Conditioned Medium of Adipose Tissue-Derived Mesenchymal Stem Cells (ATMSC)-Treated Fresh Frozen Plasma
Dian Ratih LaksmitawatiDOI: 10.12659/MSMBR.933726
Med Sci Monit Basic Res 2022; 28:e933726






Abstract
BACKGROUND: Inflammation is the body’s first response to an illness that causes irritation or infection. Inflammation is tightly correlated with aging, which is a progressive degenerative process. Conditioned medium (CM) from adipose tissue-derived mesenchymal stem cells (CM-ATMSCs) has been shown to stimulate collagen synthesis and dermal fibroblast migration, as well as reduce wrinkles and improve wound healing. This study aimed to observe the production of inflammatory modulators – interleukin (IL)-1α, IL-6, IL-10, and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) – in CM-ATMSCs treated with fresh frozen plasma (FFP) at passages 3 (P3), 7, 11, and 15.
MATERIAL AND METHODS: ATMSCs P3 were obtained from liposuction of female donors, and the CM from ATMSCs was collected. Measurement of these cytokines was performed with ELISA.
RESULTS: At many passages, IL-6, a proinflammatory modulator, was discovered to be the most powerful modulator among FFP- and non-FFP-treated cells. However, CM-ATMSCs treated with FFP and in the late passage have significant differences (P<0.05) compared to non-FFP treatments and in other passages in their effects on secretion of inflammatory modulators.
CONCLUSIONS: In conclusion, CM-ATMSC has the potential to secrete proinflammatory modulators.
Keywords: Inflammation, Inflammation Mediators, Mesenchymal Stem Cells, Adipose Tissue, Culture Media, Conditioned, Female, Humans, Plasma
Background
Skin aging is a gradual degenerative process triggered by several factors, such as UV irradiation, smoke, and stress [1]. Wrinkles are one of the characteristics of photoaging, which is associated with inflammatory response and oxidative stress that cause degradation of collagen fibrils and gelatin fibers [2]. Inflammation is the body’s first response to an illness that causes irritation or infection [3]. Interleukin (IL)-1β and IL-6 and tumor necrosis factor α (TNF-α) are the proinflammatory mediators released by macrophages and circulated throughout the body [4,5]. Other’s macrophage products such as reactive nitrogen species, reactive oxygen species, prostaglandin, and nitric oxide play essential roles in inflammation. An excessive amount of proinflammatory cytokines can also result in severe inflammation and tissue necrosis [6]. Nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) is a protein complex that controls cytokine production, and it plays an essential role in inflammation and immunity [7]. NF-κB activity is responsible for the cellular stress response, which contributes to aging [8].
Use of mesenchymal stem cells (MSCs) is one of the potential alternatives in the therapy of various diseases. MSCs from human adipose tissue (hATMSC) are known to be an attractive source of autologous MSC therapy because of the easy and repeatable access to adipose tissue [9]. Previous study shows that MSC can inhibit the proliferation of T and B cells, the production of H2O2 from neutrophils, the secretion of immunoglobulins, and the cytotoxicity of T and natural killer cells. Moreover, conditioned medium (CM) ATMSC has been reported to upregulate the secretion of some inflammatory cytokines such as TNF-α, IL-1α, IL-1β, IL-10, IL-6, and IL-4 by stimulating the resident receptor and modulating the immune response [10,11].
Fetal bovine serum (FBS) is used as a standard and reference in culture media, but it must be replaced to avoid any drawbacks such as cell interaction with animal chemicals. Several previous studies showed that use of human blood-derived serum has advantages over using fetal bovine serum (FBS) [12]. Therefore, this study aimed to determine the concentration of inflammatory modulators IL-1α, IL-10, IL-6, and NF-κB in passages 3 (P3), 7 (P7), 11 (P11), and 15 (P15) of ATMSC and then to compare their concentrations in a different medium (fresh frozen plasma [FFP] and non-FFP).
Material and Methods
ATMSC PREPARATION:
Experiments were conducted on ATMSCs P3 that were obtained from liposuction of female donors in accordance with the Institutional Ethics Committee of the Faculty of Medicine, Padjadjaran University, Bandung, Indonesia (No. 1062/UN6.C1.3.2/KEPK/PN/2016). ATMSCs from P3 were seeded at a density of 104 cells/cm2 on plastic-surfaced culture disks with 80% MEM-α, 20% FFP, 1% antibiotic and antimycotic, and 1% heparin and incubated in a humidified, 37°C, 5% CO2 environment. Cells were detached using 0.25% trypsin EDTA solution when cultures reached 80% confluence (Gibco, 25200072). P3 detached cells were grown in a separate flask until confluence was reached. In brief, the medium was collected and centrifuged at 1600 rpm for 5 min at a room temperature of 20°C, and the supernatant was filtered through a 0.22-μm filter unit (TPP, 99722) and kept at −80°C as CM-ATMSCs [13].
CHARACTERIZATION OF ATMSCS:
The ATMSCs were cultured at a density of 2×106 cells (P3) and were harvested after reaching 80% confluence. The characterization of MSCs was performed by flow cytometry (Analyzer 10, MACSQuant; Miltenyi Biotec, Germany) using specific antibodies (CD90 FIT C, CD105 PerCP-Cy5, CD73 APC, CD34 PE, CD116 PE, CD19 PE, CD45 PE, HLA-DR PE, and CD44 PE) according to the manufacturer’s procedure (BD Stemflow™ kit, 562245; BD Biosciences, NJ, USA). The surface marker studies and measurements were done in triplicate [14,15].
MEASUREMENT OF INFLAMMATORY IMMUNOMODULATOR CONCENTRATION IN CM-ATMSCS (IL-1α, IL-6, IL-10, NF-κβ):
Measurement of each inflammatory modulator was performed with ELISA using an ELISA Kit (H1587, H1386, H0483, H0088, and R0015, respectively; Elabscience, Wuhan, China). Sample and standard working solutions were prepared at approximately 100 μL in each well with 1×106 cells. Then, the solution was incubated at 37°C for 90 min. The liquid in each well was removed, and the biotinylated detection Ab working solution was added up to 100 mL. The solution was mixed thoroughly before being incubated for 60 min at 37°C. Following incubation, the solution was aspirated from each well, and 350 μL of wash buffer was added and then soaked for 1–2 min. This step was repeated 3 times.
The solution was then added with as much as 100 μL of horseradish peroxidase conjugate working solution. The solution was incubated for another 30 min at 37°C. Then, the solution was washed 5 times with 350 μL of wash buffer. The procedure was continued with the addition of 90 μL of substrate reagent and closed with a new sealer (then incubated until the color changed into blue) and incubated for 15 min at 37°C (protected from light). Subsequently, 50 μL of stop solution was added (the solution changed color to yellow). The absorbance of the solution was measured with a wavelength of 450 nm [16–19].
STATISTICAL ANALYSIS:
Statistical analysis was performed using SPSS version 16.0 software (SPSS, Inc., Chicago, IL, USA). The analysis was evaluated by one-way AVOVA, followed by Tukey’s post hoc test, and was considered to be significant (
Results
MSCs should have lineage marker-positive CD44, CD73, CD90, and CD105 and lineage marker-negative CD11b, CD19, CD34, CD45, and HLA-DR. Fluorescein staining phycoerythrin and fluorescein isothiocyanate were used to detect ATMSC surface markers (Figure 1). FFP supplementation was found to be effective in increasing proliferation, but had no effect on characteristics of MSCs. The ATMSC characterizations are shown in Figure 1. ATMSCs from P3 exhibited characteristics of MSCs. Moreover, Figure 1 shows that P3 FFP still had MSC characteristics, as the P3 population was 99.57% CD90-, 99.52% CD105-, 99.74% CD73-, and 99.32% CD44-positive.
Samples used were treated with FFP and non-FFP at various phases. Inflammatory modulators that were observed included interleukin (IL)-1α, IL-1β, IL-6, IL-10, and NF-κB. Figure 2 shows the concentration level of each sample treated with FFP at P3 (Figure 2A), P7 (Figure 2B), P11 (Figure 2C), and P15 (Figure 2D). Based on the figure, the inflammatory modulator that had the highest concentration level in each passage was IL-6. In P3, the IL-6 concentration was 786.16 pg/mL (Figure 2A), whereas NF-κB had the lowest concentration level, with 14.20 pg/mL. At P7, the IL-6 concentration was 797.75 pg/mL (Figure 2B), and IL-1α had the lowest concentration, with 17.53 pg/mL. At P11, the IL-6 concentration was 843.97 pg/mL (Figure 2C), and NF-κB had the lowest concentration, with 55.59 pg/mL. At P15, the IL-6 concentration was 871.96 pg/mL (Figure 2D), whereas IL-1α had the lowest concentration, with 23.33 pg/mL. It is also shown that IL-6 had the highest concentration at P15.
Figure 2 also shows the concentration level of each sample treated with non-FFP. According to the figure, IL-6 had the highest concentration when compared to the other modulators. At P3 (Figure 2A), the IL-6 concentration was 597.13 pg/mL, whereas NF-κB had the lowest concentration, with 15.42 pg/mL. At P7 (Figure 2B), the IL-6 concentration was 705.41 pg/mL, whereas IL-1α had the lowest concentration, with 19.67 pg/mL. At P11 (Figure 2C), the IL-6 concentration was 725.35 pg/mL, whereas IL-1α had the lowest concentration, with 22.01 pg/mL. At P15 (Figure 2D), the IL-6 concentration was 777.26 pg/mL, whereas IL-1α had the lowest concentration, with 23.69 pg/mL. IL-6 had the highest concentration at P15. This result is in line with FFP-treated ATMSCs. Furthermore, the IL-6 concentrations at P3, P7, P11, and P15 in FFP-treated ATMSC were significantly different compared to non-FFP-treated ATMSCs. It is also shown that IL-6 was higher in FFP-treated than in non-FFP-treated ATMSCs.
Discussion
Currently, MSCs are important and are being researched for the development of cell therapeutics in regenerative medicine. Its immunomodulatory properties, as well as its ability to secrete various trophic factors, demonstrate remarkable therapeutic effects in various preclinical disease models [20]. Cytokines and growth factors secreted by ATMSC in both FFP-supplemented and non-FFP-supplemented media exhibit inflammatory properties.
In the present study, ATMSCs at P3 to P15 were positive for the human mesenchymal stromal cell markers CD90, CD105, CD44, and CD73 (>95%) (Figure 1). The present results agree with other studies, in which Wharton’s jelly-derived mesenchymal stem cell (WJ-MSCs) were highly expressed in CD44, CD90, CD73, and CD105 at passages 4 and 8 [15,21]. ATMSCs were also positive for CD73, CD90, and CD105 and negative for CD11b or CD14, CD19, CD34, CD45, and HLA-DR on their cell surface [22].
IL-6 had the highest concentration of inflammatory cytokine found in non-FFP-supplemented media. In this study, compared to IL-6 secretion in FFP-supplemented media, IL-6 in non-FFP-supplemented media was significantly different. The concentration found in FFP-supplemented media was significantly higher than the concentration in non-FFP-supplemented media (Figure 2). IL-6 plays an important role in acute inflammation, and it is responsible for fever and acute-phase response in the liver [23]. In a previous study, IL-6 was produced in both FBS-supplemented and platelet-rich plasma (PRP)-supplemented media in ATMSCs [12], showing that PRP did not affect the IL-6 secretion. In another study, IL-6 was secreted and found in CM-ATMSC in high concentration but not as high as in WJ-MSC [10,12].
NF-κB showed a higher level in non-FFP-treated at late passage. CM-ATMSCs treated with FFP supplementation in the late passage (P15) can activate NF-κB signaling mediator and then upregulate the production of proinflammatory cytokines (IL-1α and IL-6) (Figure 3). This is in line with another study showing that CM-MSC stimulated NF-κB signaling, which can produce high levels of IL-6 and IL-8. This mechanism can induce the inflammation process, apoptosis, and degradative enzymes, and most probably becomes an activated aging process [24]. The activation of this protein can also promote cancer cell growth. The inhibition of NF-κB can reduce oxidative stress and disrupt senescence both in vitro and in vivo. A previous study also stated that the expression of NF-κB has a different process between young MSCs and old MSCs [25].
The results of our study are also in line with other studies, in which CM-ATMSC also produced a high level of IL-6 [25,26]. Previous studies also found that IL-6 is significantly higher in CM-MSC than in the control medium and other MSCs derived from ovarium and Wharton’s jelly [23,27,28]. However, the secretion of CM-MSCs rich in cytokines and other chemokines can also participate in wound healing by promoting cell migration, proliferation, and angiogenesis, thus neutralizing antibody against IL-6, and it can enhance the repair effect in the uterus [29]. The late passage of CM also affects the secretion of IL-6. The late passage exhibited the lowest population doubling time; therefore, it might increase the secretion of cytokines, including IL-6 [23]. The other factor that affects secretion is treatment with FFP. In another study, using platelet secretome treatment was observed to release several proteins that can act on the processes involved in wound healing and tissue repair [26].
At every passage of cells, the anti-inflammatory cytokine IL-10 was found to have a lower concentration than IL-6, and there was no significant difference in IL-10 between FFP- and non-FFP-treated cells [30]. This agrees with a previous finding showing that there was no difference in the level of IL-10 between the CM and control medium [27]. In other studies, IL-10 was not secreted by ATMSC in FBS-supplemented and PRP-supplemented media [12]. High concentrations of IL-6 and IL-10 indicate a severe infection, but an elevated IL-10 concentration does not necessarily indicate a poor prognosis [31].
In most studies, FBS is used as a standard and reference in culture media, but its substitution is required to avoid any cell contact with animal compounds. Human blood-derived usage for cell expansion in culture has been proposed in many studies. The supplementation of culture media with platelet-rich plasma (PRP) is known to induce proliferation by secreting growth factors, as well as release of extracellular matrix components and proinflammatory and angiogenic cytokines [32,33].
Conclusions
FFP supplementation and late passage have an effect on the secretion of inflammatory modulators. The older the passage, the higher the IL6 produced. Thus, the use of CM-ATMSC in anti-aging therapy should consider the passages of ATMSC to avoid high levels of proinflammatory cytokines. CM-ATMSC plays a significant role in producing proinflammatory cytokines.
Figures
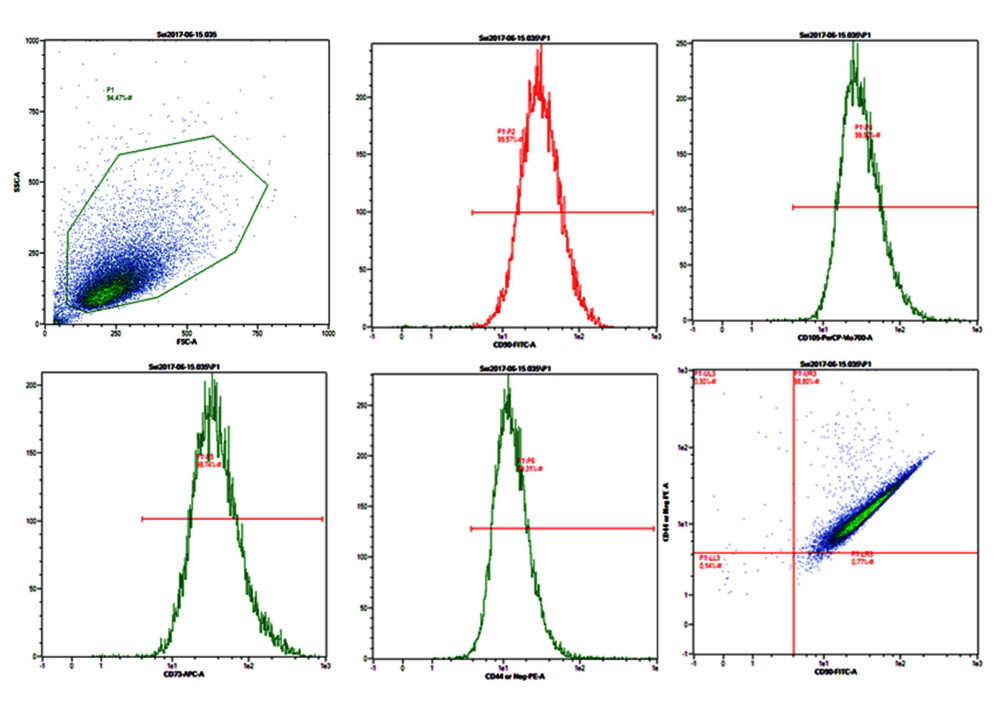 Figure 1. The percentage of surface marker ATMSCs. CD90=99.57%, CD105=99.52%, CD73=99.74%, CD44=99.32%.
Figure 1. The percentage of surface marker ATMSCs. CD90=99.57%, CD105=99.52%, CD73=99.74%, CD44=99.32%. 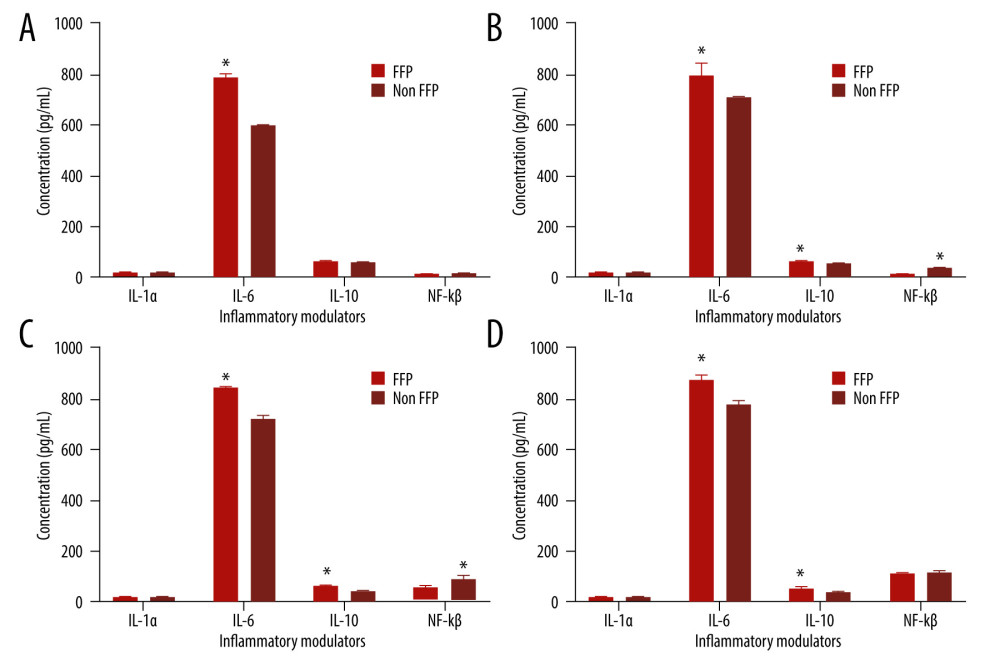 Figure 2. The concentration of inflammatory modulators (IL-1α, IL-6, IL-10, and NF-κB) treated with FFP and non-FFP at passages 3 (A), 7 (B), 11 (C), and 15 (D). * Significant difference in modulators at different treatments (P<0.05) as analyzed by independent-samples t test.
Figure 2. The concentration of inflammatory modulators (IL-1α, IL-6, IL-10, and NF-κB) treated with FFP and non-FFP at passages 3 (A), 7 (B), 11 (C), and 15 (D). * Significant difference in modulators at different treatments (P<0.05) as analyzed by independent-samples t test. 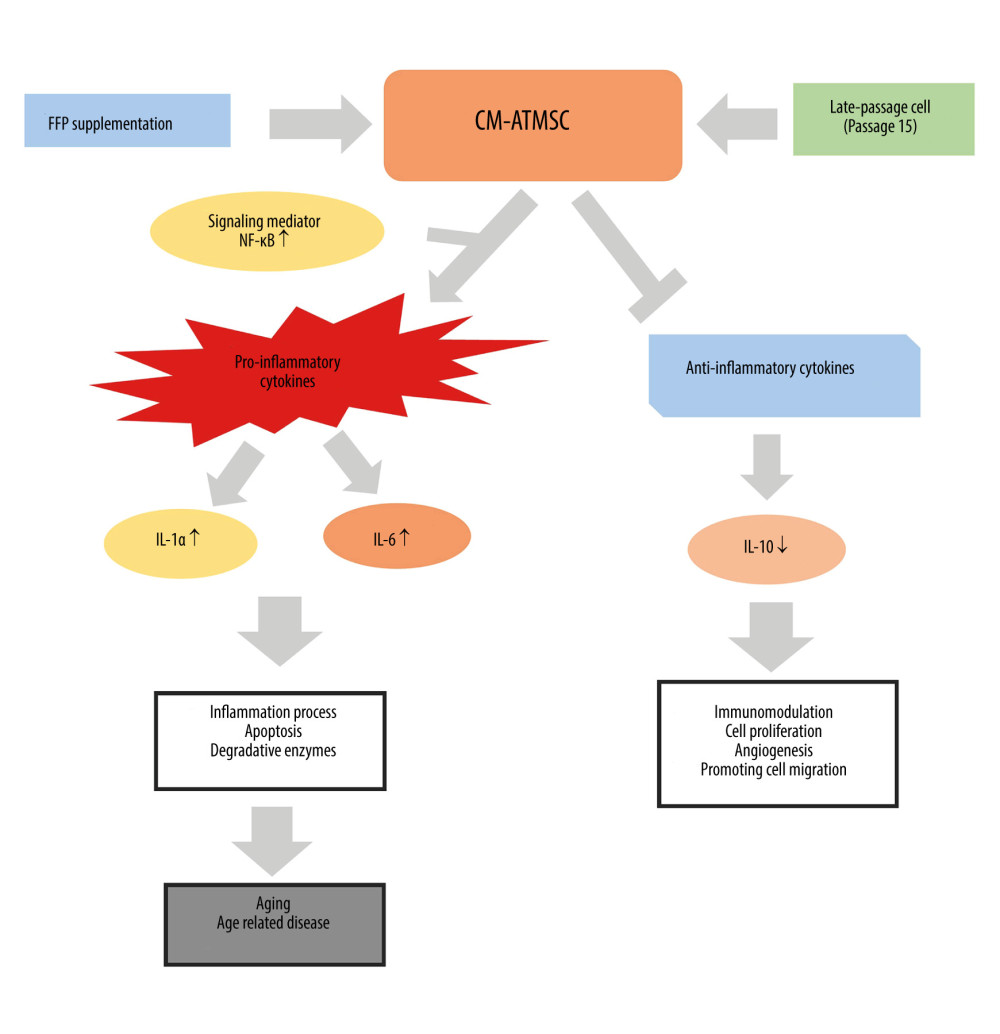 Figure 3. The potential mechanisms of CM-ATMSC with FFP supplementation treatment in an inflammation process. FFP – fresh frozen plasma; NF-κB – nuclear factor kappa-light-chain-enhancer of activated B cell; IL – interleukin.
Figure 3. The potential mechanisms of CM-ATMSC with FFP supplementation treatment in an inflammation process. FFP – fresh frozen plasma; NF-κB – nuclear factor kappa-light-chain-enhancer of activated B cell; IL – interleukin. References
1. Amani H, Shahbazi MA, D’Amico C, Microneedles for painless transdermal immunotherapeutic applications: J Control Release, 2021; 330; 185-217
2. Kim SR, Jung YR, An HJ, Anti-wrinkle and anti-inflammatory effects of active garlic components and the inhibition of MMPs via NF-κB signaling: PLoS One, 2013; 8; e73877
3. Li Q, Han SM, Song WJ, Anti-inflammatory effects of Oct4/Sox2-overexpressing human adipose tissue-derived mesenchymal stem cells: In Vivo, 2017; 31; 349-56
4. Wang WY, Tan MS, Yu JT, Role of pro-inflammatory cytokines released from microglia in Alzheimer’s disease: Ann Transl Med J, 2015; 3; 136
5. Choi SA, Lee JY, Wang KC, Human adipose tissue-derived mesenchymal stem cells: characteristics and therapeutic potential as cellular vehicles for prodrug gene therapy against brainstem gliomas: Eur J Cancer, 2011; 48; 129-37
6. Ivanova-Todorova E, Bochev I, Dimitrov R, Conditioned medium from adipose tissue-derived mesenchymal stem cells induces CD4+FOXP3+ cells and increases IL-10 secretion: J Biomed Biotechnol, 2012; 2012; 295167
7. Amable PR, Teixeira MVT, Carias RBV, Mesenchymal stromal cell proliferation, gene expression and protein production in human platelet-rich plasma-supplemented media: PLoS ONE, 2014; 9; 104662
8. Widowati W, Darsono L, Suherman J: Nat Prod Sci, 2016; 22; 147-53
9. Kakudo N, Minakata T, Mitsui T, Proliferation-promoting effect of platelet-rich plasma on human adipose-derived stem cells and human dermal fibroblasts: Plast Reconstr Surg, 2008; 122; 1352-60
10. Fong EL, Chan CK, Goodman SB, Stem cell homing in musculoskeletal injury: Biomaterials, 2011; 32; 395-409
11. Wu XB, Liu Y, Wang GH, Mesenchymal stem cells promote colorectal cancer progression through AMPK/mTOR-mediated NF-κB activation: Sci Rep, 2016; 6; 1420
12. Asumda FA, Age-associated changes in the ecological niche: implications for mesenchymal stem cell aging: Stem Cell Res Ther, 2013; 4; 1-11
13. Noverina R, Widowati W, Ayuningtyas W, Growth factors profile in conditioned medium human adipose tissue-derived mesenchymal stem cells (CM-hATMSCs): Clin Nutr, 2019; 24; 3-44
14. Widowati W, Wijaya L, Bachtiar I, Effect of oxygen tension on proliferation and characteristics of Wharton’s jelly derived mesenchymal stem cells: Biomarkers Genomic Med, 2014; 6; 43-48
15. Widowati W, Noverina R, Ayuningtyas W, Proliferation, characterization and differentiation potency of adipose tissue-derived mesenchymal stem cells (AT-MSCs) cultured in fresh frozen and non-fresh frozen plasma: Int J Mol Cell Med, 2019; 8; 283
16. Ng PC, Li K, Wong RP, Proinflammatory and anti-inflammatory cytokine responses in preterm infants with systemic infections: Arch Dis Child Fetal Neonatal Ed, 2003; 88; 1209-13
17. Widowati W, Prahastuti S, Ekayanti NLW, Anti-inflammation assay of black soybean extract and its compounds on lipopolysaccharide-induced RAW264.7 cell: J Phys Conf, 2019; 1374; 012052
18. Saanin SN, Wahyudianingsih R, Afni M: Indian J Nat Prod Resour, 2021; 11(4); 260-66
19. Laksmitawati DR, Widyastuti A, Karami N: Bangladesh J Pharmacol, 2017; 12(1); 35-40
20. Al-Sharabi N, Mustafa M, Ueda M, Conditioned medium from human bone marrow stromal cells attenuates initial inflammatory reactions in dental pulp tissue: Dent Traumatol, 2017; 33; 19-26
21. Widowati W, Wijaya L, Murti H, Conditioned medium from normoxia (WJMSCs-norCM) and hypoxia-treated WJMSCs (WJMSCs-hypoCM) in inhibiting cancer cell proliferation: Biomarkers Genomic Med, 2015; 7; 8-17
22. Corotchi MC, Popa MA, Remes A, Isolation method and xeno-free culture conditions influence multipotent differentiation capacity of human Wharton’s jelly-derived mesenchymal stem cells: Stem Cell Res Ther, 2013; 4; 81
23. D’Esposito V, Passaretti F, Perruolo G, Platelet-rich plasma increasesgrowth and motility of adipose tissue-derived mesenchymal stem cells andcontrols adipocyte secretory function: J Cell Biochem, 2015; 1; 2408-18
24. Yi PF, Bi WY, Shen HQ, Inhibitory effects of sulfated 20(S)-ginsenoside Rh2 on the release of pro-inflammatory mediators in LPS-induced RAW264.7 cells: Eur J Pharmacol, 2013; 712; 60-66
25. Gordon S, The macrophage: Past, present, and future: Eur J Immunol, 2007; 37; 9-17
26. Lawrence T, The nuclear factor NF-κB pathway in inflammation: Cold Spring Harb Perspect Biol, 2009; 1; a001651
27. Tilstra J, Clauson C, Niedernhofer L, NF-κB in aging and disease: Aging Dis, 2011; 2; 449
28. Schafer R, Spohn G, Baer P, Mesenchymal stem/stromal cells in regenerative medicine: can preconditioning strategies improve therapeutic efficacy: Transfus Med Hemother, 2016; 43; 256-67
29. Widowati W, Widyastuti H, Murti H, Interleukins and VEGF secretome of human Wharton’s jelly mesenchymal stem cells-conditioned medium (hWJMSCs-CM) in different passages and oxygen tensions: Biosci Res, 2017; 14; 776-87
30. Ding D, Liu H, Chu T, Interleukin-6 from ovarian mesenchymal stem cells promotes proliferation, sphere and colony formation and tumorigenesis of an ovarian cancer cell line SKOV3: J Cancer, 2016; 7; 1815-23
31. Ho C, Lan C, Liao C, Mesenchymal stem cells and their conditioned medium can enhance the repair of uterine defects in a rat model: J Chin Med Assoc, 2017; 81; 268-76
32. Wu Y, Hoogduijn M, Baan C, Adipose tissue-derived mesenchymal stem cells have a heterogenic cytokine secretion profile: Stem Cells Int, 2017; 2017; 4960831
33. Amable PR, Teixeira MVT, Carias RBV, Mesenchymal stromal cell proliferation, gene expression and protein production in human platelet-rich plasma-supplemented media: PloS one, 2014; 9; e104662
Figures
Figure 1. The percentage of surface marker ATMSCs. CD90=99.57%, CD105=99.52%, CD73=99.74%, CD44=99.32%.Figure 2. The concentration of inflammatory modulators (IL-1α, IL-6, IL-10, and NF-κB) treated with FFP and non-FFP at passages 3 (A), 7 (B), 11 (C), and 15 (D). * Significant difference in modulators at different treatments (P<0.05) as analyzed by independent-samples t test.Figure 3. The potential mechanisms of CM-ATMSC with FFP supplementation treatment in an inflammation process. FFP – fresh frozen plasma; NF-κB – nuclear factor kappa-light-chain-enhancer of activated B cell; IL – interleukin. Most Viewed Current Articles
15 Jun 2022 : Clinical Research
Evaluation of Apical Leakage After Root Canal Obturation with Glass Ionomer, Resin, and Zinc Oxide Eugenol ...DOI :10.12659/MSMBR.936675
Med Sci Monit Basic Res 2022; 28:e936675
07 Jul 2022 : Laboratory Research
Cytotoxicity, Apoptosis, Migration Inhibition, and Autophagy-Induced by Crude Ricin from Ricinus communis S...DOI :10.12659/MSMBR.936683
Med Sci Monit Basic Res 2022; 28:e936683
01 Jun 2022 : Laboratory Research
Comparison of Sealing Abilities Among Zinc Oxide Eugenol Root-Canal Filling Cement, Antibacterial Biocerami...DOI :10.12659/MSMBR.936319
Med Sci Monit Basic Res 2022; 28:e936319
08 Dec 2022 : Original article
Use of Estimated Glomerular Filtration Rate and Urine Albumin-to-Creatinine Ratio Based on KDIGO 2012 Guide...DOI :10.12659/MSMBR.938176
Med Sci Monit Basic Res 2022; 28:e938176