07 July 2022: Laboratory Research
Cytotoxicity, Apoptosis, Migration Inhibition, and Autophagy-Induced by Crude Ricin from Seeds in A549 Lung Cancer Cell Lines
Irma Erika Herawati12ABCDEF, Ronny Lesmana345ABDEF, Jutti LevitaDOI: 10.12659/MSMBR.936683
Med Sci Monit Basic Res 2022; 28:e936683






Abstract
BACKGROUND: Ricin protein derived from Ricinus communis seeds is known to have a high toxicity to humans and animals. Several studies revealed that ricin, belonging to ribosome inactivating protein-I, has cytotoxic properties against various types of cancer cell lines.
MATERIAL AND METHODS: Crude ricin (CR) from the seeds of R. communis was investigated for its cytotoxicity on the A549 cancer cell lines using the MTS assay, and the cell death mechanism was explored using flow cytometry and Western blot methods. The cell migration was measured using a scratch/wound-healing method and the autophagy activity was explored using Western blotting.
RESULTS: CR showed cytotoxicity against A549 cancer cell lines, with an IC₅₀ of 40.94 ppm. CR induced apoptosis and necrosis, but apoptosis occurred more frequently than necrosis. Apoptosis induced by CR was mediated by the activation of caspase-9 and caspase-3. CR inhibited cell migration in a concentration- and time-dependent manner, with the highest effect occurred at the concentration of 1.0 ppm. The autophagic experiment showed that CR inhibited autophagy in A549 lung cancer cells by decreasing Beclin-1 levels while increasing Atg5 levels in a concentration-dependent manner and CR decreased LC3-II level while increasing p62 level. Cisplatin treatment also inhibited autophagy as it exhibited the same effect on those autophagic proteins as CR.
CONCLUSIONS: Our findings suggest that CR might be a potential candidate for anticancer drugs, but further study is needed to verify its anticancer properties.
Keywords: Apoptosis, Cytotoxicity Tests, Immunologic, Ricinus, A549 cells, Animals, Antineoplastic Agents, Autophagy, Cell Line, Tumor, Humans, Lung Neoplasms, Necrosis, ricin
Background
According to the Global Cancer Statistics 2018, lung cancer is one of the most prevalent cancers in terms of incidence [1]. The rise in incidence and mortality of any type of cancer, including lung cancer, has resulted in the discovery of new drugs that are toxic to cancer cells but have no or minimum harmful effect on normal cells. However, currently used anticancer drugs have high toxicity against cancer and normal cells in other body parts [2]. As a result, new anticancer drugs that are more toxic to cancer cells but less toxic to normal cells are still being discovered through chemical synthesis or isolation of new substances from natural resources.
Numerous natural compounds have been extensively studied, and many of them have been shown to inhibit cancer cell properties by their own mechanisms of action [3]. Furthermore, many plant toxins with significant anticancer properties have been identified [4]. Considerable research has been conducted on plant toxins to discover new natural substances with high potential anticancer activity.
Ricin, one of the plant toxins, is known to be highly toxic to mammalian cells [5]. This substance is derived from the seeds of
In addition to its toxicity, ricin has been reported to have potential for the treatment of several cancer diseases, including leukemia, breast cancer [9], bladder cancer [10], cervical cancer [7], and melanoma [11]. Several investigations showed that ricin has cytotoxic activity against cancer cell lines [12], induces apoptosis [7], and inhibits protein synthesis [6,7]. The fruit extract of
Although ricin shows significant anticancer activity and potential as an anticancer agent, its high toxicity is one of the greatest limitation for the ricin applications in cancer therapy. In recent years, studies on ricin have been developed to reduce its toxicity against normal cells and to more effectively delivery it to target cancer cells by means of encapsulating it into a polymer or liposome nanocarrier [7,14]. Meanwhile, research on ricin to explore its cytotoxicity against other cancer cells and also to verify its mechanism of action continues to be carried out.
To date, the cytotoxicity profiles of ricin in lung cancer cells have not been widely reported. This study reports the cytotoxicity, apoptotic activity, cell migration inhibition, and autophagic activity of crude ricin (CR) against A549 lung cancer cell lines.
Lung cancer has a high level of inherent resistance and frequently develops treatment resistance during the course of the disease [15]. Chemotherapy and radiation resistance can be caused in part by faulty apoptotic signaling [16]. Apoptosis, a programmed cell death mechanism, plays an important role during lung cancer progression. Some morphological aspects of apoptosis include plasma membrane blebbing, cell shrinkage, chromatin condensation/fragmentation, and cell breakdown into apoptotic bodies. All of these properties are the result of caspases, a group of cystein-aspartate enzymes, selectively proteolyzing proteins important in cell signaling, DNA repair, and structural DNA integrity. Caspases are divided into 2 groups: initiator caspases (caspase-2, −8, −9, and −10 in human cells), which cleave and activate the second group, the effector caspases (caspase-3, −6, and −7 in human cells), which subsequently undertake selective proteolysis [17]. One way of increasing apoptotic signaling to improve chemotherapy and radiotherapy responses is to reactivate death receptor signaling [18,19]. In contrast, necrosis is associated with abnormal processes, induced by stimuli outside the cell or tissue, such as toxins, various infections, trauma, and ischemia, which cause death of cells prematurely [20]. Necrosis is virtually always harmful and can be lethal, while apoptosis has several advantages for the organism.
Autophagic activity is of interest to study because it plays an essential role in cell survival and death [21]. It inhibits cell survival and induces cell death, leading to the suppression of tumor formation, but it also promotes tumor formation by promoting cancer cell growth and tumor growth [22]. In addition, it has been reported that some cytotoxic drugs induce protective autophagy in cancer cells, resulting in inhibition of the drug’s cytotoxicity and apoptotic activity [22].
Material and Methods
PLANT MATERIALS:
EXTRACTION:
The extraction procedure used to obtain CR from R. communis seeds was described in our previous report [23] and followed the method of Kumar et al (2004) [24]. Briefly, the seeds were defatted by grinding them in ether and centrifuging them at 3000 rpm for 10 min. Then, the residue was resuspended in ether after discarding the supernatant. This procedure was repeated 4 times. The final residue was collected and air-dried, and then dry fat-free residue was soaked in 5-volume distilled water at pH 4.0 with the addition of dilute acetic acid. The suspension was homogenized and left for 24 h. The homogenate was further centrifuged at 8000 g for 10 min, and the supernatant was collected, adjusted to 60% ammonium sulfate saturation, set aside for 24 h at 4°C, and centrifuged for 30 min. Finally, the pellet was collected and dissolved in a mixture of 0.005 M of Na3PO4 and 0.2 M NaCl buffer pH 7.2 phosphate buffer saline (PBS). The dissolved pellet was concentrated by lyophilization and denoted as CR. The presence of ricin protein in CR was identified by employing liquid chromatography, column liquid chromatography, and fast protein liquid chromatography, followed by SDS-PAGE [23].
CELL CULTURE AND TREATMENT:
A549 lung cancer cell lines were purchased from the American Type Culture Collection ATCC CCL-185 (Manassas, VA, USA). The cell lines were cultured in RPMI-1640 medium (Sigma-Aldrich, St. Louis, MO, USA) supplemented with 10% fetal bovine serum (Sigma-Aldrich) and antibiotics, which consisted of 100 units/mL penicillin and 100 μg/mL streptomycin (Sigma-Aldrich). The cells were cultured in a standard culture condition with 5% CO2 and 37°C.
CYTOTOXICITY EXAMINATION BY 3-(4,5-DIMETHYLTHIAZOL-2-YL)-5-(3-CARBOXYMETHOXYPHENYL)-2-(4-SULFOPHENYL)-2H-TETRAZOLIUM ASSAY:
Cell cytotoxic analysis was performed using a 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS) assay on cells in the presence of various concentrations of ricin from
FLOW CYTOMETRIC ANALYSIS:
A549 cells were seeded in a 6-well plate at a density of 5×105 cells/well and were cultured for 24 h at 37 °C with 5% CO2. The cultures were treated with 0.5 ppm, 1 ppm, and 2 ppm of cisplatin and CR and incubated for 24 h at 37°C with 5% CO2. After 24 h of treatment, cells were observed by an inverted phase-contrast microscope (Olympus Corporation) at ×100 and ×200 magnification. The cells were harvested using 0.01% trypsin at 37°C for 5 min. The cells were collected in 2-mL tubes and washed twice with cold PBS via centrifugation for 5 min at 300 rpm at room temperature. The cells were gently resuspended in 1× binding buffer at a concentration of 1×106 cells/mL. Further analysis was performed using the FITC-Annexin V apoptosis detection kit I (BD Pharmingen; BD Biosciences), according to the manufacturer’s protocol. Briefly, 100 μL of the sample solution (1×105 cells/mL) was transferred to a 5-mL culture tube. A total of 5 μL FITC-Annexin V and 5μL PI was added to the tube for 15 min in the dark at room temperature. Finally, 400 μL binding buffer was added to the tubes. The samples were analyzed immediately within 1 h by using a flow cytometer (Beckton Dickinson Facslyric) and were analyzed using Flowjo software version 10.5.3 [25].
CELL EXTRACTION AND WESTERN BLOT:
One million A549 cells cultured for 24 h, after reaching 70% to 80% confluence, were treated with CR (25 ppm and 50 ppm) for 36 h. A549 cells incubated for 36 h were obtained together with control cells that had not been treated. Cells were washed twice with cold PBS buffer before being undergoing lysis with lysis buffer; protein cells were counted using the BSA method. Equally, 40 μg of cell extract was electrophoresed on a 30% polyacrylamide gel (Invitrogen, Carlsbad, CA, USA) and electro-transferred to an Amersham Protran 0.45-μm NC (GE Healthcare Life Sciences, Little Chalfont, UK). Lysis of cells was performed using RIPA lysis buffer. Immunoblotting was then performed as per the manufacturer’s guidelines (Bio-Rad, Hercules, CA, USA) using the protein extraction reagent RIPA lysis buffer (20 mM Tris-HCl (pH 7.5); 150 mM NaCl; 1 mM Na2EDTA; 1 mM EGTA; 1% NP-40; 1% sodium deoxycholate; 2.5 mM sodium pyrophosphate; 1 mM β-glycerophosphate; 1 mM Na3VO4; 1 μg/mL leupeptin) with added protease inhibitors and 1 mM sodium orthovanadate (a phosphatase inhibitor). The tissue lysates were separated by centrifugation for 15 min at 15 000 rpm and 4°C. Protein concentrations in the supernatant were measured through Bradford protein assay (Bio-Rad). Equal amounts of protein were resolved by SDS-PAGE, then transferred to a nitrocellulose membrane and immunoblotted with the antibody. Apoptosis-related proteins were analyzed by immunoblot analysis using caspase-3 (R&D Systems, Inc, Minneapolis, MN, USA) and caspase-9 antibodies (Santa Cruz Biotechnology, Inc, Dallas, TX, USA) at a 1: 1000 dilution. Autophagy signaling was analyzed by immunoblot analysis by Beclin-1 (R&D) 1: 1000 dilution; ATG5 (R&D) 1: 1000 dilution; LC3 (Cell Signaling Technology) 1: 500 dilution; and p62 (SQSTM1) (Cell Signaling Technology) 1: 1000 dilution in PBST and were incubated for 24 h. β-actin (Santa Cruz Biotechnology) served as the loading control. The membranes were imaged (LI-COR Odyssey Clx Western Blot Imager, USA), and the intensity of the bands was quantified using LiCOR quantification software [26].
MIGRATION ASSAY:
In the scratch/wound-healing assay, cell migration was observed in A549 lung cancer cell lines with a 24-well Transwell plate system (Costar, Corning, USA). The cell lines were grown in 24-well plates and treated or untreated with CR and cisplatin (a positive control) in the complete medium, and then placed in an incubator. After 24 h incubation, gaps were introduced with a P10 pipette tip by gently scraping the monolayer. After scraping, cells were washed 3 times with PBS to remove debris. Cells with gaps were incubated for 0 h, 24 h, and 48 h at 37 °C (5% CO2). The treatments were observed with a microscope linked to a computer and Toupview software (version ×64, 3.7.7892) and documented as TIFF, and the gap area was measured with ImageJ (NIH). The experiments were carried out in triplicate [27].
STATISTICAL ANALYSIS:
Data were analyzed using 1-way analysis of variance (ANOVA) using GraphPad Prism version 9.0 software for Windows. The data are presented as the mean±standard error of the mean (SEM
Results
CYTOTOXICITY OF CR ON A549 LUNG CANCER CELL LINES:
CR and cisplatin (a standard cancer drug) were examined for their cytotoxicity on A549 lung cancer cells using the MTS method with a PrestoBlue reagent (Thermo Fisher). The effects of CR and cisplatin at various concentrations (1.5, 5.0, 15, 50, 150, 500, and 1500 ppm) on the A549 cell death after 48 h were evaluated to determine their IC50 values (Figure 1). CR showed cytotoxicity with an IC50 value of 40.94 ppm, higher than the IC50 of cisplatin, which was 10.98 ppm. The cytotoxicity of CR on the lung A549 cancer cells was also higher than that on normal (MRC5) lung cells (unpublished data).
APOPTOTIC ACTIVITY IN FLOW CYTOMETRY ANALYSIS:
Apoptotic activity of CR (0.5, 1, and 2 ppm) and cisplatin (0.5, 1, and 2 ppm) was tested against A549 cancer cell lines using a flow cytometry method (Beckton Dickinson Facslyric) with Annexin V-FITC/PI reagent. All data obtained were analyzed by Flowjo software version 10.5.3. In Figure 2, the colors shown represent cells that absorb fluorescence against Annexin or PI reagents, as recorded by the ultraviolet light. Cisplatin and CR show the fractions of viable (Q4), early apoptotic (Q3), late apoptotic (Q2), and necrotic (Q1) A549 cancer cells.
The viability percentage of cisplatin- and CR-treated A549 cancer cells at every concentration was lower than that of the control (96.85%). The viability percentage of living cells of the CR-treated cells decreased in a concentration-dependent manner and was less than that of the cisplatin-treated cells. CR exhibited apoptotic activity in a concentration-dependent manner that was observed in early and late apoptosis. The cells undergoing apoptosis were found more often in the early phase of apoptosis than in the late phase. The earliest apoptotic fraction of the A549 cells (38.35%) was found at the concentration of 2 ppm of CR. Furthermore, the number of cells undergoing necrosis was much lower than apoptotic cells, indicating that CR may prefer to induce apoptosis rather than necrosis. When compared to cisplatin, CR had stronger apoptotic activity, with the number of apoptotic cells induced being 2.16-fold higher than that with cisplatin (Table 1).
CR INCREASED APOPTOTIC SIGNALING:
The caspase-inducing activity of CR and cisplatin was examined on A549 lung cancer cell lines by a Western blot method, and the results are shown in Figure 3. CR and cisplatin increased the expression levels of active fragments of caspase-9 and caspase-3 at 36 h, including caspase-9 fragments of 47 kDa and 35 kDa and caspase-3 fragments of 35 kDa and 19kDa. In this study, we found a significant increase in caspase-9 levels (Figure 3A, 3B), 2.32-fold at a dose of 25 ppm and a 3.25-fold at 50 ppm compared with the negative control. There was also a 1.46-fold and 1.84-fold increase in cleaved caspase-9 (Figure 3A, 3B) after CR treatment at doses of 25 ppm and 50 ppm, respectively. Caspase-3 protein levels were likewise enhanced by CR (Figure 3A, 3B) by 3.02-fold at 25 ppm, and 3.81-fold at 50 ppm. Meanwhile, protein levels of cleaved caspase-3 increased (Figure 3A 3B) 1.30-fold and 1.24-fold after the CR treatment at doses of 25 ppm and 50 ppm, respectively. These results suggested that the molecular mechanism of A549 lung cancer cell death caused by CR and cisplatin occurred through the activation of caspase-9 and caspase-3.
CR INHIBITED CELL MIGRATION IN THE A549 CELL LINE:
CR (0.1, 0.5, and 1.0 ppm) and cisplatin (0.1, 0.5, and 1.0 ppm) were evaluated for their inhibition against A549 cell migration after 24- and 48-h incubation by a wound-healing assay. CR and cisplatin inhibited cell migration in a concentration- and time-dependent manner, as shown in Figure 4A and 4B. The highest inhibition of both CR and cisplatin was demonstrated by the concentration of 1.0 ppm with gap enclosures of 3.10% and 3.28% in the 48-h incubation, respectively (Table 2).
CR DECREASED AUTOPHAGY SIGNALING:
The effect of cisplatin and CR on autophagy in A549 cells is shown in Figure 5A and 5B. The cisplatin treatment decreased Beclin-1 and increased Atg5 levels, and the CR treatment also decreased the Beclin-1 level 0.61-fold and 0.44-fold at doses of 25 ppm and 50 ppm, while increasing the Atg5 level by 1.32-fold and 1.45-fold at doses of 25 ppm and 50 ppm, respectively, compared with the negative control, in a concentration-dependent manner. We also performed Western blot analysis, which revealed that the CR treatment decreased LC3-II levels 0.97-fold and 0.68-fold at doses of 25 ppm and 50 ppm, respectively, while LC3-I levels decreased by 0.77-fold and 0.67-fold at doses of 25 ppm and 50 ppm, respectively. Meanwhile, the protein level of p62 increased 1.78-fold and 7.76-fold after the CR treatment at 25 ppm and 50 ppm. As p62 is known to bind LC3, it serves as a selective autophagic substrate. These results suggest that CR and cisplatin inhibited autophagy in A549 lung cancer.
Discussion
Ricin, known as a highly toxic substance derived from the seeds of
Further experiments were conducted to determine whether the cytotoxicity of CR on the A549 cancer cells was related to the induction of apoptotic cell death, so the apoptotic activity of CR was evaluated using the flow cytometric method (Figure 2), followed by the Western blot method (Figure 3). The flow cytometric analysis indicated that cisplatin and CR significantly decreased the number of living cells compared with the control (Figure 2). Furthermore, the viability percentage of living cells in the CR-treated cells decreased in a concentration-dependent manner. The cell death caused by CR was due to apoptosis as well as necrosis, but apoptosis occurred more frequently than necrosis, indicating that CR may be more potent in inducing apoptosis than necrosis (Table 1). Moreover, apoptosis may occur rapidly because it is seen that most apoptosis occurs in the early stage rather than the late stage.
Analysis of the mechanism of apoptotic activity of CR on the caspase cascade pathway in the Western blot method indicated that CR induced apoptosis intrinsically via a mitochondrial caspase-9 signaling pathway and activation of caspase-3 (Figure 3). It signifies that the cell death was caused by the induction of apoptosis through the activation of caspase-9 as the initiator caspase and caspase-3 as the effector caspase.
In cancer therapy, anticancer drugs are generally expected not only to suppress cancer cell growth but also to prevent cell metastases. The migration and invasion of cancer cells into different tissues is an important initial step in metastases. These processes can be targeted by an anticancer agent to inhibit cancer cell development and aggression. In this study, the effect of CR and cisplatin on A549 cancer cell migration was examined by the scratch/wound assay [29], which demonstrated that CR and cisplatin inhibited cell migration in a concentration- and time-dependent manner as compared with the control (Figure 4). The highest inhibition of both agents was provided by the concentration of 1.0 ppm with the gap enclosure of 3.10% and 3.28% within the 48-h incubation, respectively (Table 2). To date, no report has been found concerning the inhibitory activity of ricin protein against A549 cancer cell migration, and the only evidence reported is the inhibition of the fruit extract of
Chemotherapy is a common strategy for cancer treatment, but its success rate is often limited by the development of chemotherapy resistance in different cancer cells to multiple anticancer drugs [22]. This is due, in particular, to the induction of drug-protected autophagy bypassing the apoptotic pathway [30].
In the present study, CR inhibited autophagic responses in A549 cancer cells. The inhibition of autophagy was found through a Western blot analysis of autophagic-related proteins. CR decreased LC3-II expression while increasing p62 expression (Figure 5). The LC3-II is known as the only protein biomarker reliably associated with autophagosome formation and maturation, and p62 is a binding protein serving as a link between LC3 and substrate of autophagy [31]. The decreased LC3-II and increased p62 levels indicate that autophagosome formation was inhibited. This was also accompanied by a decrease of Beclin-1 and an increase of Atg5, which are involved in the regulation of autophagy. Beclin-1 is a core component of the class III phosphatidylinositol-3-kinase/Vps34 complex I, which is essential for the phagophore nucleation step of autophagy [32]. Atg5 is an important protein involved in the expansion of isolation membranes and is required to form autophagosomes [31]. This evidence suggests that CR may inhibit rather than induce autophagy in A549 human lung cancer. It is of considerable interest to find evidence that CR can induce apoptosis and inhibit autophagy because chemotherapeutic drugs commonly induce apoptosis as well as protective autophagy, which contribute to chemotherapeutic resistance in many types of cancer cells. As a result, the discovery of chemotherapeutic agents having apoptotic activity and inhibiting effects on autophagy might be an important target for many researchers.
Many studies have shown the potential of ricin as an anticancer agent; however, for the application in cancer therapy, ricin has many limitations, such as non-specificity, immunogenicity, fast degradation in the blood stream, and lysosomal degradation inside cells [7]. Our present findings concerning the cytotoxic activity of CR against lung cancer cells with some molecular mechanism profiles have provided additional evidence of ricin’s anticancer potential. This new evidence resulted from our efforts to find out more specific target cells for ricin. The characteristics of cancer cells may affect the specificity of an anticancer agent to interact with cancer cells. When the lung cancer cells can have more sensitive and specific characteristics to ricin, it is very possible for ricin to interact more effectively with target cells and less effectively on normal cells. This discovery is expected to reduce the limitations of ricin in terms of non-specificity. However, the data obtained are still incomplete, and much work needs to be done to provide the significance of the therapeutic application of ricin.
This study revealed that cisplatin inhibited autophagy in A549 lung cancer cells after 36 h of treatment, as evidenced by decreased levels of Beclin-1 and LC3-II and increased levels of Atg5 and p62. These findings contradicted previous evidence that cisplatin induces protective autophagy in many types of cancer cells, such as ovarian cancer [33], H460 cells [34], oesophageal cancer [35], and bladder cancer [36]. However, it is in line with the results reported by Bello et al (2013) [32] that autophagy decreases below the basal level of control cells in cisplatin-treated melanoma cells. This was further reduced in still-living adherent and floating cells after 24 h of treatment, when significant apoptosis has occurred. After 18 to 20 h of treatment, and in a shorter period of time, there was no substantial change in LC3-II levels, indicating that the length of the incubation can affect the results.
Conclusions
Taken together, the results of this study provided evidence that CR has cytotoxicity against A549 lung cancer cells with the apoptotic death mechanism through the activation of caspase-9 and caspase-3. Moreover, CR inhibited A549 cell migration and autophagy in a concentration- and time-dependent manner. These findings suggest that CR might have potential as a novel anticancer drug candidate. However, the limitation of ricin concerning its high toxicity to human cells is still a great challenge to overcome. Further studies on the effect of isolated ricin on these particular A549 lung cancer cells are needed to verify its anticancer properties.
Figures
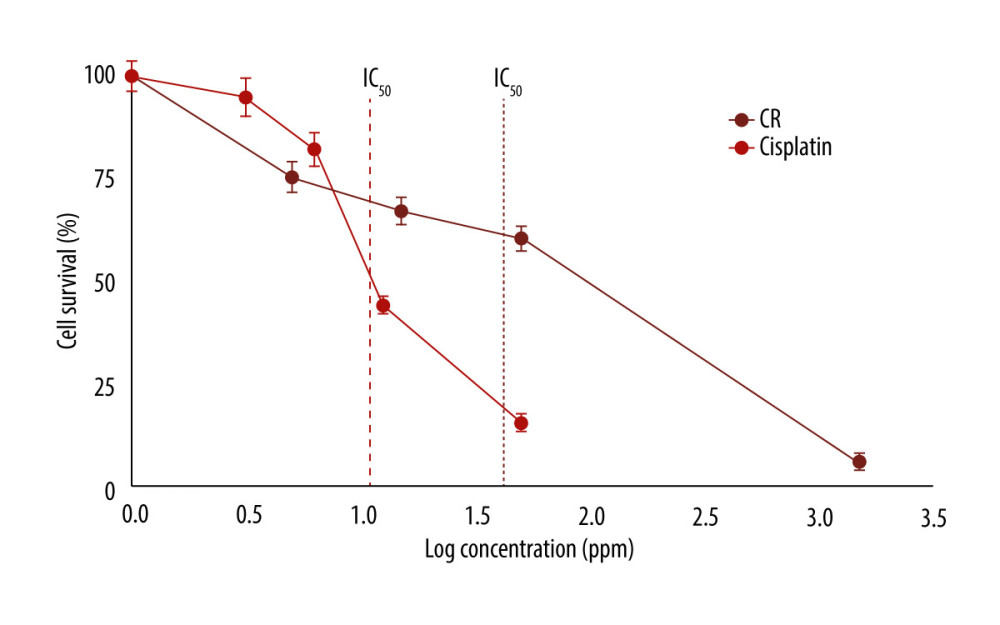 Figure 1. Cytotoxicity of crude ricin (CR) and cisplatin on A549 lung cancer cells at various concentrations in the 48-h treatment. Data are presented as mean±standard error minimum (SEM) (n=3).
Figure 1. Cytotoxicity of crude ricin (CR) and cisplatin on A549 lung cancer cells at various concentrations in the 48-h treatment. Data are presented as mean±standard error minimum (SEM) (n=3). 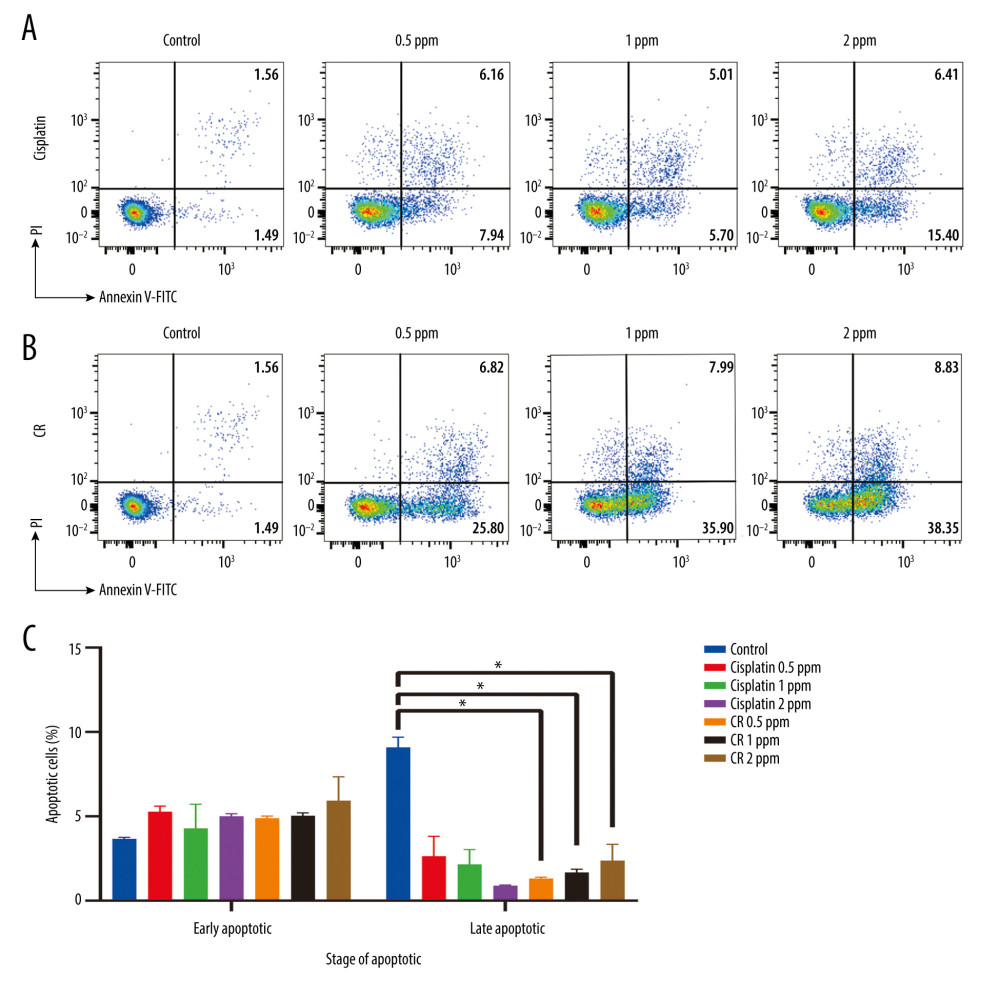 Figure 2. Annexin V/PI double-staining assay of A549 cells treated with 0.5 ppm, 1 ppm, and 2 ppm of cisplatin and crude ricin (CR) for 24 h. Representative flow cytometry result showing (A) cisplatin-dose-dependent and (B) CR dose-dependent alteration distribution of early and late apoptosis. (A1) and (B1) show negative control of cell line, (A2–A4) show increased apoptotic cells after cisplatin treatment, (B2–B4) show increased apoptotic cells after CR treatment. (C) Bar graph shows the average increased proportion of early and late apoptosis after cisplatin and CR treatment. Data are presented as mean±standard error minimum (SEM) (n=3). One-way ANOVA followed by Tukey’s post-hoc test was conducted. * P<0.05.
Figure 2. Annexin V/PI double-staining assay of A549 cells treated with 0.5 ppm, 1 ppm, and 2 ppm of cisplatin and crude ricin (CR) for 24 h. Representative flow cytometry result showing (A) cisplatin-dose-dependent and (B) CR dose-dependent alteration distribution of early and late apoptosis. (A1) and (B1) show negative control of cell line, (A2–A4) show increased apoptotic cells after cisplatin treatment, (B2–B4) show increased apoptotic cells after CR treatment. (C) Bar graph shows the average increased proportion of early and late apoptosis after cisplatin and CR treatment. Data are presented as mean±standard error minimum (SEM) (n=3). One-way ANOVA followed by Tukey’s post-hoc test was conducted. * P<0.05. 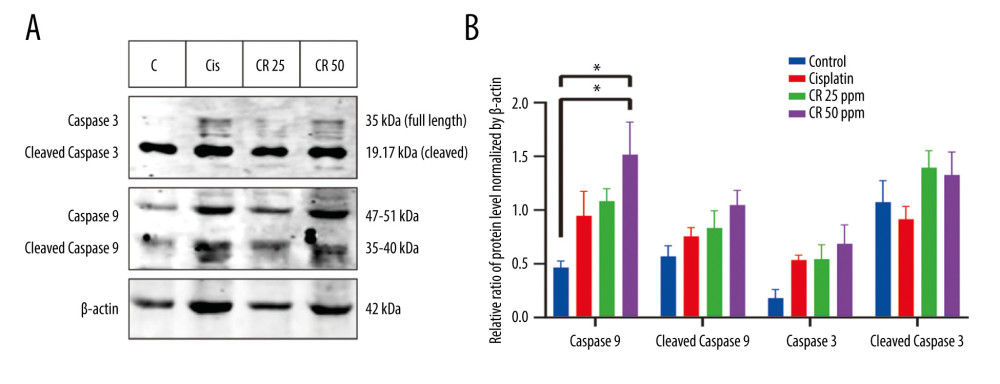 Figure 3. Western blot analysis of caspase cascade-related proteins in A549 lung cells treated with cisplatin and CR. (A) Representative blots of caspase-9 and -3 were determined in A549 cells treated with cisplatin and CR at 36 h; (B) Densitometry protein levels of caspase-9, cleaved caspase-9, caspase-3, and cleaved caspase-3, measured using Image Studio Digits v. 5.2. and presented as a bar graph. The data are presented as mean±standard error minimum (SEM) (n=3). A one-way ANOVA followed by Tukey’s post-hoc test was conducted. * P<0.05.
Figure 3. Western blot analysis of caspase cascade-related proteins in A549 lung cells treated with cisplatin and CR. (A) Representative blots of caspase-9 and -3 were determined in A549 cells treated with cisplatin and CR at 36 h; (B) Densitometry protein levels of caspase-9, cleaved caspase-9, caspase-3, and cleaved caspase-3, measured using Image Studio Digits v. 5.2. and presented as a bar graph. The data are presented as mean±standard error minimum (SEM) (n=3). A one-way ANOVA followed by Tukey’s post-hoc test was conducted. * P<0.05. 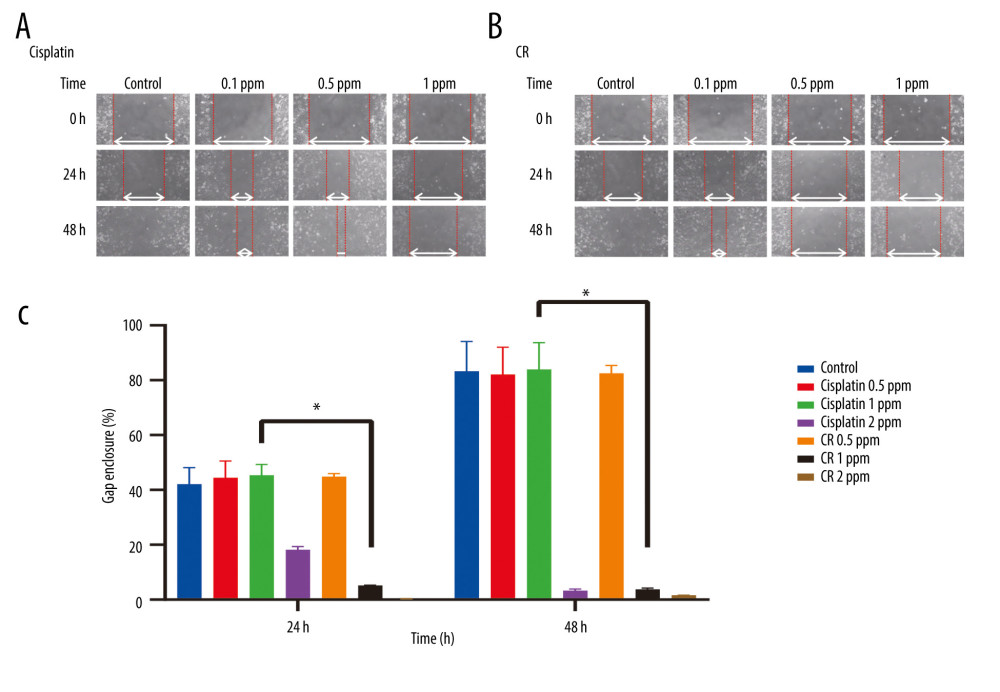 Figure 4. Effect of cisplatin and crude ricin (CR) on migration activity of A549 cell lines. (A) and (B) are representative photographs of cell migration of A549 cell lines treated with cisplatin and CR for 24 h and 48 h, respectively. The white line reflects gap enclosure. (C) Quantification gap enclosure (% area). Data are presented as mean±standard error minimum (SEM) (n=3). One-way ANOVA followed by a Tukey’s post-hoc test was conducted. * P<0.05.
Figure 4. Effect of cisplatin and crude ricin (CR) on migration activity of A549 cell lines. (A) and (B) are representative photographs of cell migration of A549 cell lines treated with cisplatin and CR for 24 h and 48 h, respectively. The white line reflects gap enclosure. (C) Quantification gap enclosure (% area). Data are presented as mean±standard error minimum (SEM) (n=3). One-way ANOVA followed by a Tukey’s post-hoc test was conducted. * P<0.05. 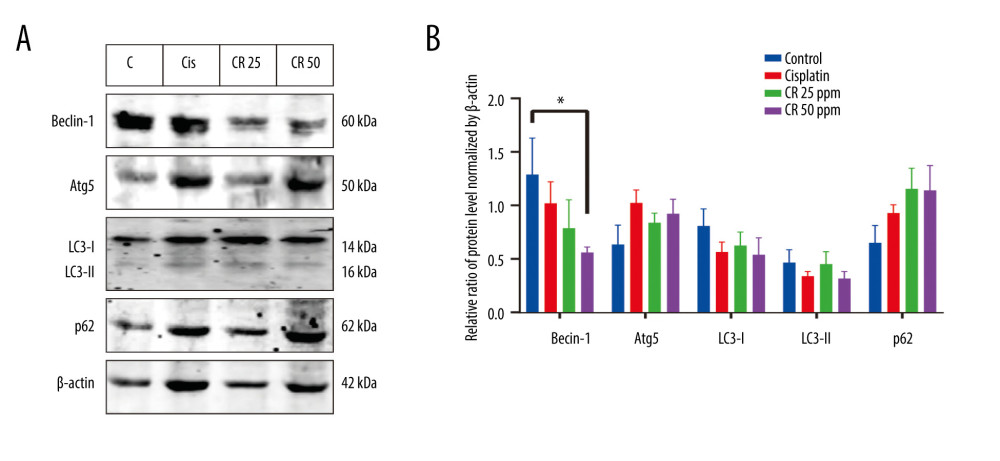 Figure 5. (A) Representative blot of Beclin-1, Atg5, LC3-I, LC3-II, and p62 determined in A549 cells treated with cisplatin and crude ricin (CR) at 36 h; (B) densitometry protein levels of Beclin-1, Atg5, LC3-I, LC3-II, and p62 measured using Image Studio Digits version 5.2 and presented as a bar graph. The data are presented as mean±standard error minimum (SEM) (n=3). One-way ANOVA was used, followed by Tukey’s post-hoc test. * P<0.05.
Figure 5. (A) Representative blot of Beclin-1, Atg5, LC3-I, LC3-II, and p62 determined in A549 cells treated with cisplatin and crude ricin (CR) at 36 h; (B) densitometry protein levels of Beclin-1, Atg5, LC3-I, LC3-II, and p62 measured using Image Studio Digits version 5.2 and presented as a bar graph. The data are presented as mean±standard error minimum (SEM) (n=3). One-way ANOVA was used, followed by Tukey’s post-hoc test. * P<0.05. Tables
Table 1. Percentage of viable, early apoptotic, late apoptotic, and necrotic A549 cancer cells in the cisplatin and crude ricin (CR) treatment with significant effect between control and CR in stage of apoptosis (a,b,c).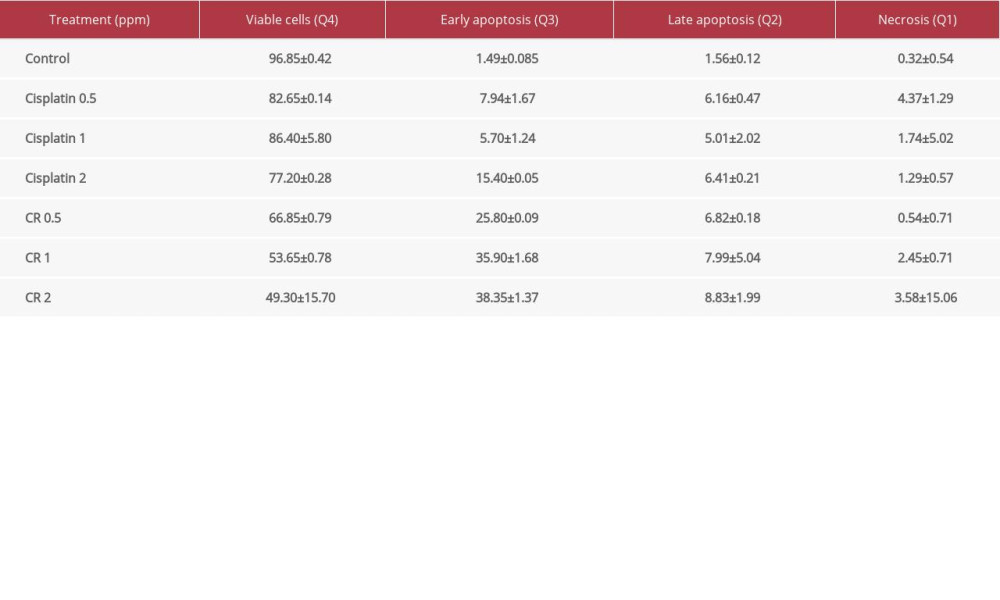 Table 2. Percentage of gap enclosure induced by cisplatin and crude ricin (CR) in A549 lung cancer cell lines.
Table 2. Percentage of gap enclosure induced by cisplatin and crude ricin (CR) in A549 lung cancer cell lines.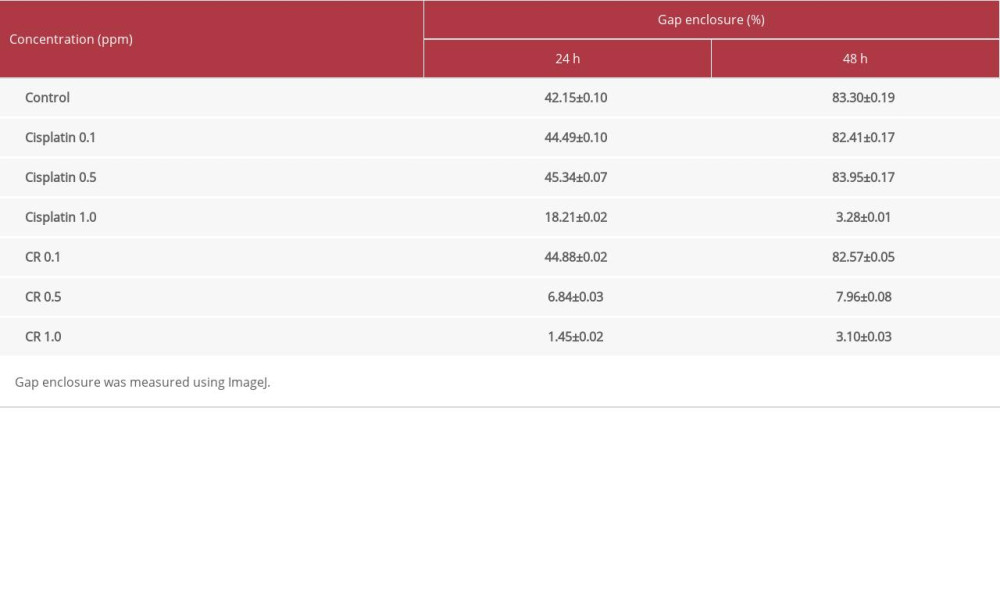
References
1. Bray F, Ferlay J, Soeriomataram I, Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries: Cancer J Clin, 2018; 68(6); 394-424
2. Sakarkar DM, Deshmukh VN, Ethnopharmacological review of traditional medicinal plants for anticancer activity: Int J Pharm Tech Res, 2011; 3; 298-308
3. Kinghorn AD, Farnsworth NR, Soejarto DD, Novel strategies for the discovery of plant derived anticancer agents: Pharm Biol, 2003; 41(Suppl 1); S53-67
4. Ram VJ, Kumari S, Natural products of plant origin as anticancer agents: Drug News Perspect, 2001; 14; 465-82
5. Lord JM, Roberts LM, Robertus JD, Ricin: Structure, mode of action, and some current applications: FASEB J, 1994; 8; 201-8
6. Rogozinska NS, Sominka H, Golacka JN, Intracellular transport and cytotoxicity of the protein toxin ricin: Toxins, 2019; 11(6); 350
7. Tyagi N, Tyagi M, Pachauri M, Potential therapeutic applications of plant toxin-ricin in cancer: Challenges and advances: Tumor Biol, 2015; 36(11); 8239-46
8. Polito L, Bortolotti M, Battelli MG, Ricin: An ancient story for a timeless plant toxin: Toxins (Basel), 2019; 11; 324
9. Tyagi N, Rathore SS, Ghosh PC, Efficacy of liposomal Monensin on the enhancement of the antitumor activity of liposomal Ricin in human epidermoid carcinoma (KB) cells: Indian J Pharm Sci, 2013; 75; 16-22
10. Li C, Yan R, Yang Z, BCMab1-Ra, a novel immunotoxin that BCMab1 antibody coupled to Ricin A chain, can eliminate bladder tumor: Oncotarget, 2017; 8(28); 46704-5
11. Spitler LE, Del RM, Khentigan A, Therapy of patients with malignant melanoma using a monoclonal antimelanoma antibody-ricin A chain immunotoxin: Cancer Res, 1987; 47; 1717-23
12. Trung NN, Tho NT, Dung BTT, Effects of ricin extracted from seeds of the castor bean (ricinus communis) on cytotoxicity and tumorigenesis of melanoma cells: Biomed Res Ther, 2016; 5(3); 633-44
13. Majumder M, Debnath S, Gajbhiye RL: Sci Rep, 2019; 9; 14493
14. Epler K, Padilla D, Phillips G, Delivery of ricin toxin a-chain by peptide-targeted mesoporous silica nanoparticle-supported lipid bilayers: Adv Health Mater, 2012; 1; 348-53
15. Joseph B, Lewensohn R, Zhivotovsky B, Role of apoptosis in the response of lung carcinomas to anti-cancer treatment: Ann N Y Acad Sci, 2000; 926; 204-16
16. Johnstone RW, Ruefli AA, Lowe SW, Apoptosis: A link between cancer genetics and chemotherapy: Cell, 2002; 108; 153-64
17. Nicholson DW, Caspase structure, proteolytic substrates, and function during apoptotic cell death: Cell Death Different, 1999; 6; 1028-42
18. Kelley SK, Ashkenazi A, Targeting death receptors in cancer with Apo2L/TRAIL: Curr Opin Pharmacol, 2004; 4; 333-39
19. Cretney E, Takeda K, Smyth MJ, Cancer: Novel therapeutic strategies that exploit the TNF-related apoptosis-inducing ligand (TRAIL)/TRAIL receptor pathway: Int J Biochem Cell Biol, 2007; 39; 280-86
20. Proskuryakov SY, Konoplyannikov AG, Gabai VL, Necrosis: A specific form of programmed cell death?: Exp Cell Res, 2003; 28(1); 1-16
21. Levine B, Autophagy and cancer: Nature, 2007; 446; 745-47
22. Yun CW, Lee SH, Review: The roles of autophagy in cancer: Int J Mol Sci, 2018; 19; 3466
23. Herawati IE, Lesmana R, Levita J, Analysis of crude ricin from Ricinus communis originated from Nganjuk, East Java, Indonesia, using liquid chromatography, column liquid chromatography, and fast protein liquid chromatography (FPLC): Rasayan J Chem, 2022; 15(1); 204-9
24. Kumar O, Nashikkar A, Jayaraj R, Purification and biochemical characterisation of ricin from castor seeds: Defence Sci J, 2004; 54(3); 345-51
25. Amalia E, Diantini A, Subarnas A, Water-soluble propolis and bee pollen of Trigona spp. from South Sulawesi Indonesia induce apoptosis in the human breast cancer MCF-7 cell line: Oncol Lett, 2020; 20; 274
26. Setiawan I, Lesmana R, Goenawan H, Calcitriol potentially alters HeLa cell viability via inhibition of autophagy: J Cancer Res Therapeut, 2021 [Ahead of print publication]
27. Schneider CA, Rasband WS, Eliceiri KW, NIH image to imageJ: 25 years of image analysis: Nat Methods, 2012; 9(7); 671-75
28. Derbyshire EJ, Henry RV, Stahel RA, Potent cytotoxic action of the immunotoxin SWAll-ricinA chain against human small cell lung cancer cell lines: Br J Cancer, 1992; 66; 444-451
29. Cory G, Scratch-wound assay. Cell migration. Methods in molecular biology (methods and protocols): Methods Mol Biol, 2011; 769; 25-30
30. Kondo Y, Kanzawa T, Sawaya R, The role of autophagy in cancer development and response to therapy: Nat Rev Cancer, 2005; 25(9); 726-34
31. Klionsky DJ, Abdelmohsen K, Abe A, Guidelines for the use and interpretation of assays for monitoring autophagy: Autophagy, 2016; 12(1); 1-222
32. Bello BD, Toscano M, Moretti D, Cisplatin-induced apoptosis inhibits autophagy, which acts as a pro-survival mechanism in human melanoma cells: PLoS One, 2013; 8(2); e57236
33. Wang J, Wu GS, Role of autophagy in cisplatin resistance in ovarian cancer cells: J Biol Chem, 2014; 289(24); 17163-73
34. Sirichanchuen B, Pengsuparp T, Chanvorachote P, Long-term cisplatin exposure impairs autophagy and causes cisplatin resistance in human lung cancer cells: Mol Cell Biochem, 2012; 364(1–2); 11-18
35. O’Donovan TR, O’Sullivan GC, McKenna SL, Induction of autophagy by drug-resistant esophageal cancer cells promotes their survival and recovery following treatment with chemotherapeutics: Autophagy, 2011; 7(5); 509-24
36. Lin JF, Lin YCh, Tsai TF, Cisplatin induces protective autophagy through activation of BECN1 in human bladder cancer cells: Drug Des Devel Ther, 2017; 11; 1517-33
Figures
Figure 1. Cytotoxicity of crude ricin (CR) and cisplatin on A549 lung cancer cells at various concentrations in the 48-h treatment. Data are presented as mean±standard error minimum (SEM) (n=3).Figure 2. Annexin V/PI double-staining assay of A549 cells treated with 0.5 ppm, 1 ppm, and 2 ppm of cisplatin and crude ricin (CR) for 24 h. Representative flow cytometry result showing (A) cisplatin-dose-dependent and (B) CR dose-dependent alteration distribution of early and late apoptosis. (A1) and (B1) show negative control of cell line, (A2–A4) show increased apoptotic cells after cisplatin treatment, (B2–B4) show increased apoptotic cells after CR treatment. (C) Bar graph shows the average increased proportion of early and late apoptosis after cisplatin and CR treatment. Data are presented as mean±standard error minimum (SEM) (n=3). One-way ANOVA followed by Tukey’s post-hoc test was conducted. * P<0.05.Figure 3. Western blot analysis of caspase cascade-related proteins in A549 lung cells treated with cisplatin and CR. (A) Representative blots of caspase-9 and -3 were determined in A549 cells treated with cisplatin and CR at 36 h; (B) Densitometry protein levels of caspase-9, cleaved caspase-9, caspase-3, and cleaved caspase-3, measured using Image Studio Digits v. 5.2. and presented as a bar graph. The data are presented as mean±standard error minimum (SEM) (n=3). A one-way ANOVA followed by Tukey’s post-hoc test was conducted. * P<0.05.Figure 4. Effect of cisplatin and crude ricin (CR) on migration activity of A549 cell lines. (A) and (B) are representative photographs of cell migration of A549 cell lines treated with cisplatin and CR for 24 h and 48 h, respectively. The white line reflects gap enclosure. (C) Quantification gap enclosure (% area). Data are presented as mean±standard error minimum (SEM) (n=3). One-way ANOVA followed by a Tukey’s post-hoc test was conducted. * P<0.05.Figure 5. (A) Representative blot of Beclin-1, Atg5, LC3-I, LC3-II, and p62 determined in A549 cells treated with cisplatin and crude ricin (CR) at 36 h; (B) densitometry protein levels of Beclin-1, Atg5, LC3-I, LC3-II, and p62 measured using Image Studio Digits version 5.2 and presented as a bar graph. The data are presented as mean±standard error minimum (SEM) (n=3). One-way ANOVA was used, followed by Tukey’s post-hoc test. * P<0.05. Tables
Table 1. Percentage of viable, early apoptotic, late apoptotic, and necrotic A549 cancer cells in the cisplatin and crude ricin (CR) treatment with significant effect between control and CR in stage of apoptosis (a,b,c).Table 2. Percentage of gap enclosure induced by cisplatin and crude ricin (CR) in A549 lung cancer cell lines.Table 1. Percentage of viable, early apoptotic, late apoptotic, and necrotic A549 cancer cells in the cisplatin and crude ricin (CR) treatment with significant effect between control and CR in stage of apoptosis (a,b,c).Table 2. Percentage of gap enclosure induced by cisplatin and crude ricin (CR) in A549 lung cancer cell lines. Most Viewed Current Articles
13 Apr 2020 : Original article 22,361
Outcome of 24 Weeks of Combined Schroth and Pilates Exercises on Cobb Angle, Angle of Trunk Rotation, Chest...DOI :10.12659/MSMBR.920449
Med Sci Monit Basic Res 2020; 26:e920449
20 Apr 2018 : Original article 19,876
Brain Training Games Enhance Cognitive Function in Healthy SubjectsDOI :10.12659/MSMBR.909022
Med Sci Monit Basic Res 2018; 24:63-69
23 Jul 2016 : Review article 12,190
Cardiac Hypertrophy: An Introduction to Molecular and Cellular BasisDOI :10.12659/MSMBR.900437
Med Sci Monit Basic Res 2016; 22:75-79
26 Sep 2024 : Original article 11,790
Methods of Processing Dental Chromium-Cobalt Alloys for Production of Metal Frameworks Faced with Ceramics ...DOI :10.12659/MSMBR.944051
Med Sci Monit Basic Res 2024; 30:e944051